Introduction
Accumulated data about the role of prolactin (PRL) in human breast cancer appear to be controversial. Thus, it has been difficult to establish definitive involvement of PRL in human breast disease. PRL was first recognizedas a hormone that plays an important role in breast cancer initiation and development in rodents (1,2) and, at least partly, in humans. There is also firm evidence of a direct stimulatory role of PRL on mammary epithelial cells (3,4), and breast cancer cells in culture (5,6). There are also some epidemiological studies showing significant increase in serum PRL concentrations in certain subpopulations of breast cancer patients (7-10), and in women at risk of developing familial breast cancer (9,11). This paper examines the basis for a claim that PRL has a contributory role during breast oncogenesis.
Actions of PRL within mammary tissues – in vitro investigations
Synthesis and secretion of PRL by breast cancer cells
Both endocrine and autocrine/paracrinesources of PRL have been recognized to exist in mammals. For many years, evidence in the literature has hinted at the possibility of an extrapituitary source of PRL. Evidence from the 1970’s indicatedthat hypophysectomized breast cancer patients had near-normalPRL levels (12,13), whereas immunohistochemistry studies revealedthe expression of immunoreactive PRL protein in human breastepithelium (14). In addition, low levels of circulating PRL were found to persist in patients under pituitary hormone suppression therapy (15). Studies in the early 1990’s indicated that the mRNA for PRL couldbe found in normal and neoplastic human breast epithelium (16-18), and mammary epithelium from pregnant rodents (19,20). PRL is generated by tumors as well as by a variety of normal tissues. Placenta is the richest source of the extrapituitary PRL (21), and is responsible for its high level in human amniotic fluid. Immune system, uterus, brain and dermal fibroblasts also produce PRL (21-24). These findings led both Vonderhaar and Clevenger laboratories (17,18) to hypothesize and subsequently prove that PRLis synthesized and secreted in human breast tissues and cells. Bioactive PRL is synthesized by human breast cancer cells in culture and acts in an autocrine/paracrine stimulatory loop within breast tissue, suggesting a role for this hormone in the pathogenesis of breast cancer (17,18). The growth of both T47Dco (ER-negative) and MCF-7 (ER-positive) human breast cancer cells was inhibited after treatment with monoclonal antibodies raised against human pituitary PRL (17). In addition, antisense RNA directed against the gene encoding for pituitary PRL significantly inhibited growth of T47Dco cells (25). The presence of the mRNA for PRL in T47Dco and MCF-7 was confirmed by RT-PCR (reverse transcription polymerase chain reaction) (17), and 82% of all tested breast cancer cell lines contained mRNA for PRL (25). Furthermore, the vast majority,i.e. 98% of human breast cancers synthesize PRL mRNA as detectedby in situ hybridization (26). In addition, some studies revealed that more than 75% of primary breast cancer surgical samples also contain mRNA for PRL, and, in the majority of cases, the amount of mRNA for PRL and its receptors is significantly elevated in malignant vs. the adjacent, non-malignant tissue from the same patient (16,25,27).
PRL receptors (PRLR)
The actions of PRL in the mammary gland require the presenceof its cognate cell surface receptor, the PRLR. PRLR belongs to the cytokine hematopoietic family of receptors (28,29). The members of this superfamily are single membrane-spanning receptors with three domains: an extracellular ligand binding domain, a hydrophobic transmembrane domain, and an intracellular proline-rich domain. There are at least three different isoforms of PRLR differing mainly in their cytoplasmic domain: long (90 kDa), intermediate, and short (40kDa) isoform. Long and short isoforms of PRLR are generated by differential splicing of a single gene and differ only in the length of the cytoplasmic domain (30). The intermediate form is a deletion mutant of the long form, lacking 198 aminoacids in its cytoplasmic domain. The shortest PRLR isoform, the PRL binding protein (PRLBP) was identified in human serum (31) and it represents the freely circulating extracellular domain of the PRLR (32). All three isoforms promote mitosis (33). Both normal and malignant mammary cells contain both the long and short forms of PRLR (34,35), while the intermediate form is found in Nb2 lymphoma cells and is more sensitive to PRL compared with the other two forms of PRLR (36). The intermediate form of PRLR has been detected in breast tissue samples (37), in contrast to the results from another study (18). Advances in immunohistochemistry, insitu hybridization, and RT-PCR enabled increasingly sensitive estimationof the PRLR in human breast cancer, and results of some of the studies using thesetechnologies (26,35,37,38) have revealed that the hPRLR is expressed in upto 98% of all human breast cancers. The studies examining PRLRexpression at the mRNA level have suggested an association witheither ER/PR (estrogen/progesterone receptors) expression (38) or neoplasia (37); however, studiesat the protein level have not confirmed these observations (26).The PRLR are commonly stabilized in human breast cancer due to decreased phosphorylation of residue Ser349 which, when phosphorylated, recruits the beta Trcp E3 ubiquitin ligase and facilitates PRLR degradation (39).
Neoplastic development and progression require deregulated cell proliferation,increased cellular survival, acquisition of an adequate vascularsupply, and escape from constraints on motility. Despite evidence that PRL can trigger the growth and motility of human breast cancer cells, the inability of PRL to trigger differentiation remains uncertain. Potential mechanisms include alterations in Stat5 (signal transducer and activator of transcription) levels or phosphorylation, quantitative changes in the expression of various hPRLR isoforms, or alteration in the malignant epithelial cell’s responsiveness to the basement membrane, which could indirectly impact PRLR signaling (32). Because enhanced motility is one aspect ofthe metastatic process, it has been questionedwhether PRL could serve as a chemoattractant for human breastcancer in vitro (40). PRL stimulates the cytoskeletal r-organization and motility of breast cancer cells. During PRLR signaling, Vav2 (guanine nucleotide exchange factor) becomes phosphorylated and activated, an event regulated by the serine/threonine kinase Nek3 (never in mitosis gene a-related kinase 3) which contributes to PRL-mediated breast cancer motility through mechanisms involving Rac1 (guanine nucleotide exchange factor) activation and paxillin phosphorylation (41).
PRL may also influence mammary carcinogenesis bymodulating vascularization. It was shown that hPRL itself, as wellas human GH, and placental hormones, could also stimulate formation of capillaries inthe chicken chorioallantoic membrane assay (42). In contrast,a proteolytic cleavage product of PRL, 16K-PRL, is a potentantiangiogenic agent in vivo and in vitro (42-44). ThisN-terminal cleavage product of PRL inhibited endothelial cellproliferation in response to vascular endothelial growth factor (VEGF)and basic fibroblast growth factor (BFGF) by inhibiting the Ras-Raf1-MAPK signaling pathway and increasing expression of type 1 plasminogen activatorinhibitor (42,45,46). These activities appear to be mediatedby a receptor distinct from the PRLR (47). The induction of VEGF by PRL is PRLR-, Jak2-, and MAP kinase-dependent. PRL induces VEGF expression through Erg-1 (ether-a-go-go-related gene 1 encoded K+ channels), and implicates VEGF as an intermediary of PRL-regulated angiogenesis (48).
The role of PRL in breast cancer is complicated by the fact that PRL itself is angiogenic, but proteases cleave PRL to generate vasoinhibins, a family of peptides that act on endothelial cells to suppress angiogenesis and vasodilation and to promote apoptosis-mediated vascular regression (49).
PRL/PRLR signaling pathways
Upon PRL-induced receptor dimerization, several different kinases are activated to transduce the hormonal signal. Proximal PRLR signaling is initiated by three tyrosine kinases, namely Jak2 (Janus kinase), Src (sarcoma), and Tec (protein tyrosine kinase) (32). These kinases belong to the Janus family of kinases (Jak family). Multiple additional downstream pathways, suchas Src family kinases, Ras-MAPKs (rat sacoma-mitogen-activated protein kinases), and PI3K (phosphoinositide 3 kinase) are involved in PRL signaling (50) (Figure 1).
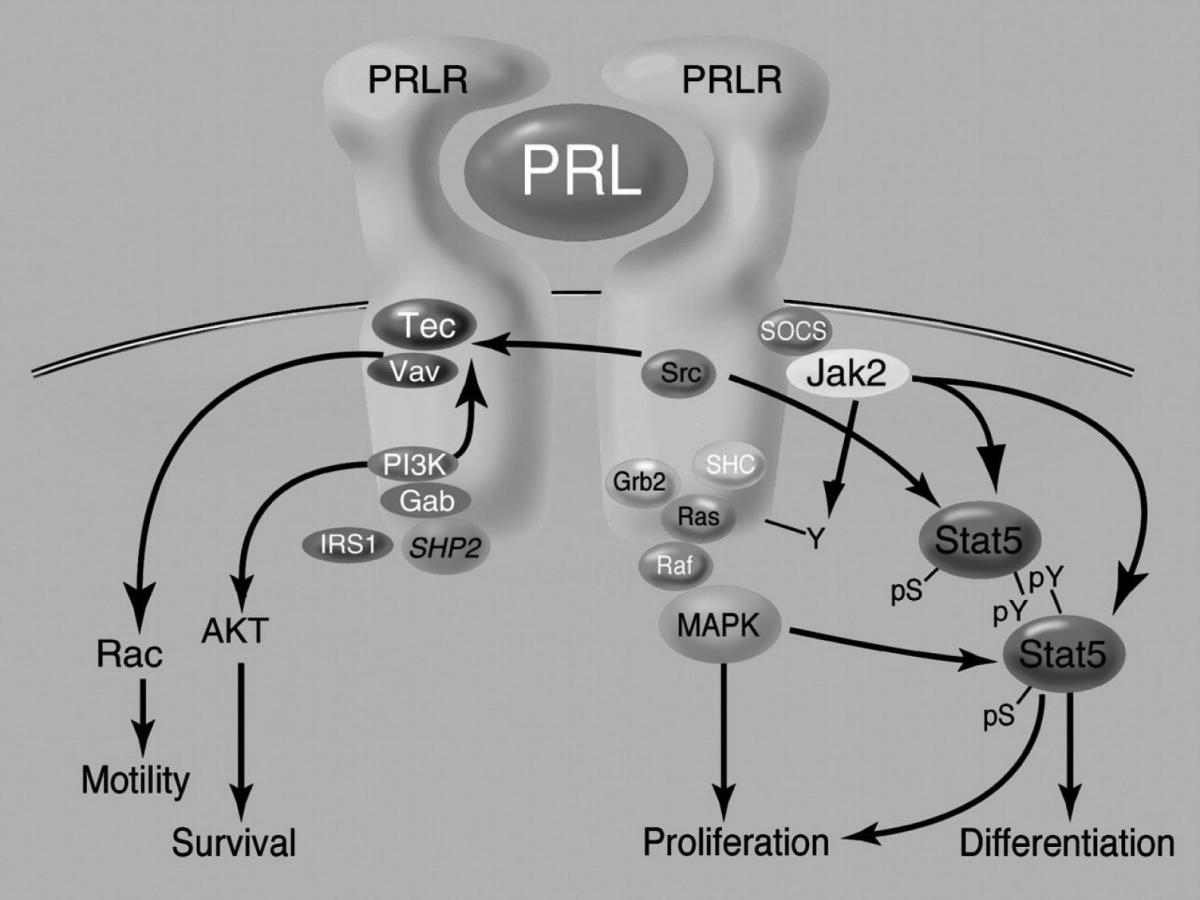
Figure 1. Aspects of PRLR signaling pathways. PRL-induced receptor dimerization induces the association of the Jak2 kinase, resulting in the activation of Jak2, PRLR phosphorylation, and the association and phosphorylation of Stat5. Signaling through the SHC/GRB2/Ras/Raf/MEK/MAPK pathway also directly stimulates proliferation and modulates Stat activity. The complex between the Tec tyrosine kinase and the Vav family of guanine nucleotide exchange factors also inducibly associates with ligand-bound PRLR resulting in its activation and stimulation of cellular motility (modified according to ref. 32).
Although it is not clear that all PRL signaling requires Jak2 as a proximal intermediate(50,51), a great deal of evidence in many cell typessupports a key role for this kinase in many actions of PRL (52,53). Jak2 phosphorylates multiple substrates, including thePRLR and Jak2 themselves. This provides docking sites for proteinswith SH2 (src homology2) domains, including Stats. The binding of PRL to its receptors induces tyrosine phosphorylation of cytoplasmic transcription factors, mainly Stat family members (32,54). Activation of Stat proteins results in their translocation to the nucleus, and subsequent activation of gene transcription (32,55). Stat1, Stat3 and Stat5 are activated in T47D cells within 15 min of PRL treatment (56). Several studies have demonstrated increased levels of Stats1 and 3 in primary mammary tumors(57,58). However, theirtarget genes in oncogenic processes, the relative importanceof PRL in their regulation, and differences from the normalmammary gland have not been understood. Both Stats3 and 5 are involvedin PRL activation of the cyclin D1 promoter (59), suggestingat least one target of PRL through this pathway that could contributeto tumorigenesis. It has beendemonstrated (60) that Stat5b, rather than Stat5a, is a potentmediator of Src-induced tumorigenesis. As discussed above, tyrosine phosphorylationby a receptor-associated Jak2 kinase resultsin the dimerization/multimerization and nuclear retrotranslocationof the Stat complex where it engages its cognate DNA bindingsequence, resulting in promoter transactivation under appropriateconditions (32,61). Whereas PRL stimulated tyrosinephosphorylation and nuclear translocation of both Stats5a and5b, Src activation resulted in tyrosine phosphorylation of bothStats5a and 5b, but nuclear translocation of only Stat5b (62).PRLhas been shown to activate Src in a variety of cell types, includingrat liver (63). Results from some recent studies indicate that proto-oncogene c-Myc potentiates Stat5a-driven gene expression, possibly functioning as a Stat5a coactivator in human breast cancer (54,64). Furthermore, PRL stimulates ubiquitination, internalization, and degradation of its receptors via catalytic activation of Jak2 (65), and it seems that Ser349 phosphorylation of PRLR is essential event in this complex Jak2-mediated signaling.
Ras-Raf-MAPK pathway
Ras-Raf-MAPK signaling pathway is a mechanism by which a variety of growth factors and cytokines mediate their proliferation action. Raf1 (mitogen activated protein kinase kinase kinase), MEK (mitogen activated protein kinase kinase), and MAP (mitogen activated protein kinase) are downstream kinases in this signaling pathway. Signaling through the SHC/GRB2/Ras/Raf/MEK/MAPK pathway (SHC-adaptor protein; GRB2-growth factor receptor-binding protein2; Ras-rat sarcoma) also directly stimulates proliferation and modulates Stat activity. PRL has been shownto activate this pathway in a number of PRL-dependent models(66), and mammary tumor cell lines (67-69), as wellas normal mouse mammary epithelial cells (68,69). PRL can also synergistically activate this pathwayvia cross-talk with other growth factors, depending on the phenotypeof the tumor cell. PRL-induced activation of Jak2 resulted intyrosine phosphorylation of erbB2, thereby increasing associationwith Grb2, and activating the Ras-MAPK pathway (70) including phosphorylation of some transcriptionfactors and thus increasing synthesis of the products of the fos gene family. Cross-talk betweenthe Stat and MAPK pathways at other points has been well documentedfor many cytokines, including PRL (71). MAPKs are able to phosphorylateStats on serine and threonine residues, which augments the activitiesof Stats1 and 3 (72).
PI3K (phosphoinositide 3 kinase) pathways
The complex between the Tec tyrosine kinase and the Vav family of guanine nucleotide exchange factors also inducibly associates with ligand-bound PRLR. This leads to the exchange of GDP for GTP on the small G protein Ras, resulting in its activation and stimulation of cellular motility. Activation of tyrosine kinases Tec and Akt (PKB-protein kinase B) is directly linked to the PRL-induced activation of PI3K. The phosphatase SHP-2 also binds to PRLR and potentiates its activity. Activation of PI3K generates phosphoinositides that serve as second messengers and canregulate multiple pathways important in oncogenesis, includingproliferation and cytoskeletal rearrangements, as well as inhibitionof apoptosis and angiogenesis (73-75). Activation of phosphoinositide cycle results in the stimulation of PKC (protein kinase C) pathway. PI3K could potentially be activated by PRL throughmultiple additional pathways. It can be a target of Ras (76),and the p85 regulatory subunit has been shown to associate withseveral downstream effectors and adaptors of cytokine and growthfactor receptors, including Stat5, Stat3, IRS1 (insulin receptor substrate 1), Gab1 (GRB2-associated binding protein 1) and Gab2 (GRB2-associated binding protein 2), and SHP-2 (Src homology-2 domain-containing protein-tyrosine phosphatase) (77,78). PI3K-generated phosphoinositides provide docking sites for Akt, as well as its upstream kinases. Phosphoinositide metabolites may also bind to guanine nucleotide exchangefactors, including Vav, as well as to Tec, a member of a largerfamily of tyrosine kinases. A constitutive complex of Tec and Vav (79) associates withPRLR in ligand-stimulated T47D (80).
Intranuclear action of PRL
Some literature data have indicated a functional role forPRL within the nucleus (81,82). These data stand in contrastwith the classic theory that postulates that peptide hormone actionis mediated only by cell surface receptors.Recent data, however, have revealed arole for the peptidyl prolyl isomerase cyclophilin B (CypB)in the nuclear transport and function of PRL (82). A complex between PRL and CypB is found in human serum, binds to the PRLR, and is endocytosed duringreceptor internalization. The PRL/CypB complexacts within the nucleus as a transcriptional inducer by facilitating the interactionof Stat5 with DNA by inducing the release of a repressor ofStat5, namely PIAS3 (83).
Rodent models of PRL action
Prolactin’s role in rodent mammary cancer has been well documented (1,84). There is a direct correlation between drug-induced hyperprolactinemia and increased tumor growth, and hypoprolactinemia and retarded tumor growth (1,85). PRL exposure enhances the development of chemicallyinduced mammary cancers in rodents (1,86,87). Overexpression of PRL in transgenic mice with increased activationof the PRLR is sufficient to induce the formation of mammarycancers at 11–15 months of age (88,89). Exposure to estrogen with secondary increases in circulatingPRL levels is able to restore susceptibility to chemical carcinogensin parous mice (90). In summary, numerous studies point toa role for PRL in increasing receptiveness to chemical carcinogensin rodent mammary glands.
Human breast cancer and PRL – in vivo investigations
Epidemiology of PRL and human breast cancer
Several epidemiological studies have indicated that PRL mayalso function as a progression factor for human breast cancer(8,91-93). A number of studies have evaluated the association between PRLlevels and several well-confirmed breast cancer risk factorssuch as parity and age at first birth, age at menarche and menopause, family history of breast cancer, mammographic density, ethnic differences, dietary intake, medication use, and prolactinomas.
PRL levels appear to decrease,at least modestly, with each additional pregnancy (94,95). Also, noindependent association between age at first birth and PRL levelwas observed (94). Overall, no significant associations between PRL and eitherage at menarche or age at menopause were reported (94,96). In a subset of subjects at risk with family history of breast cancer, basal serum PRL levels were significantly elevated (11).However, several other studies (94-96) offered scanty data to support association of the family history of breastcancer with PRL concentration.
Increased PRL levels were observedin postmenopausal women with increased breast tissue density (94,97), suggesting a measurable influence of PRL on breast epithelial and/or stromalproliferation.
Several studies evaluated PRL levels and dietaryintake (95,98) but consistent findings areyet to be reported.
A number of medications are known to increase (e.g., oral contraceptives,reserpine, haldol, cimetidine, and the phenothiazines) or decrease(e.g., levodopa) plasma PRL levels. Long-term use oforal contraceptives increases the risk of breast cancer (99). Cimetidine also increases PRL levels, but a fewstudies published have not shown any meaningful link with breast cancer (100). Women with prolactinomas have greatly elevated PRL levels.However, only a few case reports of breast cancer in women ormen with prolactinomas (101,102) and a small cohortstudy of 67 women with prolactinomas (103) have been publishedto date. Data of one recent study suggest modestpositive association between prolactin and breast cancer riskamong predominately premenopausal women; however, further follow-upis needed to increase statistical test power for subgroup analyses (104).
Genetic variations in PRL and PRL receptor (PRLR) genes as predictors of plasma PRL levels and breast cancer risk among African-American, native Hawaiian, Japanese-American, Latin, and white women were evaluated recently in a large multiethnic cohort study (105). In this comprehensive analysis covering 59 kb of the PRL locus and 210 kb of the PRLR locus, no significant association was found between common variation in these candidate genes and breast cancer risk or plasma PRL levels. The LD (linkage disequilibrium) characterization of PRL and PRLR in this multiethnic population provides a framework for studying these genes in relation to other disease outcomes that have been associated with PRL, as well as for larger studies of plasma PRL levels.
Clinical data about the association of PRL with breast cancer
The function of PRL in the etiology and progression of human breast cancer is not yet clear, and literature data are not consistent but, are even contradictory. However, there is significant evidence that PRL may play a role in human breast cancer.
The incidence of hyperprolactinemia was significantly higher in patients with metastatic breast cancer than in patients with non-metastatic breast cancer, or with mastopathy, or with advanced solid tumors of different histology (106). Hyperprolactinemia was almost exclusively found in patients with metastatic breast cancer during the course of the disease (8,107). Hyperprolactinemia was found to be an important indicator of unfavorable prognosis in node-positive breast cancer patients (108). Results of another study indicated the possible association of hyperprolactinemia and overexpression of p53 with aggressiveness of the tumor, early disease relapse or metastases, and poor overall survival in node-negative breast cancer patients (9). In primary tumors of stage II and stage III breast cancer patients, there was a significant increase in the frequency of PRL-positive tumors upon increase in the number of involved lymph nodes (109). With increasing tumor size, a significantly increased incidence of hyperprolactinemia was observed, hyperprolactinemic patients had a significantly increased risk of developing recurrent/metastatic disease, and seventy-eight per cent of their tumors showed positive immunoreactivity with PRL antibody indicating that breast tumors produce PRL which may act as a major local growth promoter (110). Results of some other studies are in agreement with those mentioned above. Circulating levels of PRL might be very useful diagnostic and prognostic marker in breast cancer patients (111), and a valuable parameter to assess treatment efficacy in breast carcinoma patients (112). Hyperprolactinemia is an indicator of disease progression and poor prognosis in metastatic breast cancer patients (113,114). Serum levels of PRL probably directly depend on the size of primary tumor in breast cancer patients, especially in those with hyperprolactinemia, but this is not a differentiation-dependent phenomenon (115).
In contrast, a surgery-induced rise in circulating PRL was associated with prolonged disease-free survival in operable breast carcinoma patients with or without axillary metastases (92,116).
Constitutive oncogenic signaling downstream of ErbB2 and Ras stabilizes PRLR via inhibitory phosphorylation of glycogen synthase kinase 3 beta (GSK3 beta) on Ser9. Importantly, inactivation of GSK3 beta correlates with elevated levels of PRLR protein in clinical human breast cancer specimens (39).
Increased prolactin levels are often associated with decreased sexual hormone levels. Association of hyperprolactinemia with reproductive disorders, amenorrhea and irregular menstrual cycles has already been known. Hyperprolactinemia in premenopausal women causes hypogonadism manifested by infertility, oligomenorrhea or amenorrhea and less often by galactorrhea (117). Patients with menstrual disturbances had higher PRL levels than women with normal menstrual function (118). However, results of one recent study have shown that no hormonal changes in serum levels of sexual hormones were found in women with hyperprolactinemia and sexual dysfunction (119). Furthermore, there are many contradictions concerning the association of female sexual steroids with cancer. Re-evaluation of earlier results supporting the carcinogenic capacity of estrogen has exhibited many shortcomings and controversies. Recently, clinical studies on hormone replacement therapy in postmenopausal women have justified beneficial anticancer effects in several organs, even in the female breast. The newly revealed association between estrogen deficiency and oral cancer risk also means a contradiction with regard to the traditional concept of estrogen-induced cancer. However cancers of highly estrogen dependent organs such as breast, endometrium and ovary exhibit both premenopausal and postmenopausal occurrence. In spite of different epidemiological data of these two groups of cancers, the mechanism of gene regulation disorder in the background of tumor initiation cannot act through entirely opposite pathways (120). This suggests that serious estrogen deficiency in moderately estrogen sensitive organs, and even mild estrogen deficiency in highly estrogen dependent organs is enough to provoke gene regulation disorders. New findings both on smoking- and hormone related cancers might lead to the same reversal; estrogen deficiency rather than estrogen itself may provoke cancer initiation (120).
Inhibition of PRL action
A decrease in PRLR levels achieved by either pharmacologic or genetic means in human breast cancer cells dramatically reduced transformation and tumorigenic properties of these cells (121). Tamoxifen (TAM), the first line of therapy in pre- and postmenopausal ER (estrogen receptor)-positive breast cancer patients also has an anti-prolactin action (122). Antilactogenic activity of TAM results from its interaction with the antilactogen binding site (ALBS) (123) which is located on the PRL receptor. ALBS is a member of the family of high affinity membrane-associated binding sites called antiestrogen binding sites (AEBS) (124). Antiestrogens, acting through the ALBS, inhibit the growth of PRL-responsive cells even in the absence of ER (125, 126). The results of these studies indicate that ALBS is in the PRL receptor, and that TAM and other related anti-estrogens may inhibit the growth of ER-negative human breast cancer cells in vitro through this mechanism (34). These data also suggest that TAM and other related drugs that act at the level of target tissue may be clinically useful in the treatment of hyperprolactinemic breast cancer patients. However, raloxifene, a selective estrogen receptor modulator and thus a breast cancer prevention agent, had no significant effect on PRL levels in premenopausal women at high risk for developing breast cancer (127).
Conclusions
PRL is synthesized by human breast cancer cells in culture, and acts in an autocrine/paracrine stimulatory loop within breast tissue. Both normal and malignant mammary cells contain both long and short forms of PRLR. PRL stimulates the cytoskeletal re-organization and motility of breast cancer cells. PRL may also influence mammary carcinogenesis bymodulating vascularization. Prolactin’s role in rodent mammary cancer has been well documented. Several epidemiological studies have indicated that PRL mayalso function as a progression factor for human breast cancer. Hyperprolactinemia was almost exclusively found in patients with metastatic breast cancer during the course of the disease, and it was found to be an important indicator of unfavorable prognosis in node-positive breast cancer patients. Taking into account all these data it could be concluded that both endocrine and autocrine PRL are involved in human breast carcinogenesis. However, much more work is needed to understand the signalingpathways used by PRL to promote tumorigenesis in mammary cells, andinteractions of these signaling cascades and their complex regulatoryloops with different oncogenes, growth factors and hormonesimportant in mammary carcinogenesis. Furthermore, the ongoing development of PRLR-specific antagonists may yield novel therapeutic strategies in treatment of human breast cancer based on blocking PRL actions at the endocrine and autocrine/paracrinelevels.