Introduction
Eosinophil cationic protein (ECP), also known under the name ribonuclease 3 (RNase 3), is a single cationic polypeptide chain consisting of 133 amino acids, with isoelectric point being 10.8 due to a high content of basic amino acids, particularly arginine. In the protein structure, there are three potential sites for N-linked glycosylation: amino acids 57-59, 65-67, and 92-94. Due to the different content of carbohydrate subunits: sialic acid, galactose and acetylglucosamine, the molecular mass of ECP protein ranges between 16 and 22 kDa (1,2). A gene for human ECP (RNASE3) is located on chromosome 14 (q24-q31). Its size is 1.2 kb and it is made of two exons: non-coding exon 1 (67 bp) and coding exon 2 (667 bp), and of one intron (230 bp) (3). Several single nucleotide polymorphisms (SNPs) in RNASE3 have been described so far which determine the cellular content and biological activity of the ECP protein (3,4). Owing to genetic factors and posttranslational reactions, ECP is a very heterogeneous molecule (5).
ECP synthesis and secretion
ECP is present in the granules of eosinophil granulocytes together with the major basic protein (MBP), eosinophil peroxidase (EPO) and eosinophil -derived neurotoxin/ eosinophil protein X (EDN/EPX). Very small amounts of ECP can be found in neutrophil granulocytes and monocytes (6,7).
ECP synthesis in eosinophil granulocytes begins already at the stage of promyelocytes in primary granules; from the myelocyte stage, ECP is present only in the matrix of specific granules of eosinophil granulocytes (8,9). Unstimulated neutrophil granulocytes cannot synthetize ECP, but this granulocyte are able to capture ECP from the environment. However, ECP mRNA is detected in neutrophil granulocytes after cellular stimulation (10). In contrast, monocytes can synthetize ECP but their ability for synthesis is lost during macrophage differentiation (7).
ECP is excreted by activated eosinophil tissue granulocytes. The secretion of ECP from eosinophil granulocytes may be induced in two manners, i.e. as antibody-dependent (imunoglobulins, Ig; IgG, IgA), and as antibody-independent (C3 and C5 complement components) activation of eosinophil degranulation (11,12). Interleukins (IL) like IL-5 and IL-3, and the granulocyte-monocyte colony stimulating factor (GM-CSF) have positive impact on this secretion. The tissue infiltrated with eosinophil granulocytes contains two types of specific eosinophil granules: intracellular cytoplasmic eosinophil granules and intact extracellular eosinophil granules. The release of the contents from intracellular granules occurs after activation of eosinophil granulocytes. With regard to this process, two mechanisms are known: the so-called “piecemeal degranulation” denotes selective secretion via secretory vesicles while exocytosis involves a nonselective process which occurs much more rarely. Extracellular granules reach a tissue after cytolysis of eosinophilic granulocytes. Their secretory properties are under the control of numerous membrane cytokine receptors that enable selective release of individual proteins from extracellular granules. Extracellular granules have been supposed to function as entirely independent structures and to additionally enhance local inflammatory reaction (13,14). During the excretion of the ECP molecule from eosinophil granules, enzymatic deglycosylaton occurs that converts the inactive ECP form with high molecular mass into a cytotoxic variant with low molecular mass (15).
Inhibitory effect on ECP secretion has been confirmed for cyclosporine A (an immunosuppressant), dexamethason (an anti-inflammatory and immunosuppressive drug), rapamycin (an immunosuppressant), formoterol (a long-acting β2-agonist), and pemirolast (an anti-allergic drug) (16-18).
RNASE3 polymorphisms
Seven SNPs have been described so far in the ECP gene, and the distribution of individual polymorphisms differs significantly between individual populations. In Caucasian populations, three polymorphisms have been most frequently identified, i.e. ECP 434(G>C), ECP 562(G>C) and ECP -38(C>A). The activity of these polymorphisms is related to the quantity of synthesized protein and biological function so that the polymorphisms have also been associated with the development of some pathological conditions (4).
ECP 434(G>C) polymorphism is located in the RNASE3 coding region. Due to G>C substitution, the codon for the amino acid arginine is changed in the codon for threonine, which leads to the change in Arg97Thr in the ECP protein. The consequence of the change in amino acid structure is alteration of biological activity of the ECP molecule because Arg97 is important for the cytotoxic activity of ECP (19,20). The substitution of arginine at position 97 with threonine allows the formation of a new, fourth glycosylation site at position 95 (ECP95Asn), and enhanced glycosylation of ECP protein decreases its cytotoxic activity. Studies of recombinant ECP97Thr have showed that, after deglycosylation, it becomes cytotoxically active like ECP97Arg (20). Parasitary infections, asthma and malignant diseases are only some pathological conditions with ECP involvement in their pathomechanisms so that disease outcome is associated with ECP phenotype or genotype. Thus, individuals infected with Schistosoma mansoni (S. mansoni) parasite, who are at the same time carriers of the ECP 434GG cytotoxic genotype, have better primary defense reaction compared with infected individuals with other genotypes, but also develop considerably more frequently a serious and life-threatening complications like hepatic fibrosis (19). The prevalence of individual ECP 434 genotype in patients with allergic diseases demonstrates ethnic and gender variability (21). Patients with allergic asthma have 434GG (ECP97Arg) genotype considerably more often than non-allergic asthmatics. It has also been observed that the combination of atopia and 434GG (ECP97Arg) genotype considerably increases the risk for development of allergic manifestations. However, as the frequency of asthmatic patients is not increased among individuals with 434GG (ECP97Arg) genotype, the polymorphism has no predictive value for the development of asthma (22). The association of ECP 434GG genotype and the occurrence of allergic symptoms is explained by the cytotoxic activity of ECP97Arg in epithelial cells in the nose and lungs of patients with allergic rhinitis and allergic asthma (4). Also, a relationship has been described between ECP 434(G>C) polymorphism and lung function. Thus, women who were 434GG genotype carriers had decreased pulmonary function, which can be accounted for by the fact that cytotoxic activity of ECP97Arg may destroy lung epithelial cells (21).
Patients with malignant diseases have inflammatory cell infiltration around tumor tissue; the task of these cells is anti-tumor action so that tumor-associated tissue eosinophilia (TATE) is associated with better disease prognosis (23,24). However, other studies have shown that the TATE has a poor prognosis or that eosinophils have an important role in tumor progression (25-27). Under diverse stimuli the eosinophils are able to induce or inhibit the tumor destruction (27). The association between TATE and ECP polymorphisms is new insight into the role of eosinophils in malignant diseases. According to Pereira and colleagues (28), patients with oral squamous cell carcinomas who are the carriers of non-cytotoxic ECP 434CC genotype have a poor clinical outcome despite tumor-associated tissue eosinophilia. In addition, research on tumor cell lines in vitro demonstrated reduced antitumor cytotoxic capacity of recombinant ECP97Thr.
ECP 562(G>C) polymorphism is located in the 3’-untranslated region of RNASE3. The intracellular concentration of ECP was significantly lower in eosinophil granulocites from subjects with ECP 562CC genotype but a relationship has not been described between the 562(G>C) polymorphism and allergy (29). The potential relationship between the 3’-untranslated region and expression of mRNA have been described in other genes (30). A possible explanation for 562(G>C) polymorphism and cellular ECP content could be alterations in the mRNA stability.
ECP -38(C>A) polymorphism is located in the first RNASE3 intron in the region of the binding site for the globin transcription factor-1 (GATA-1) which regulates the development of eosinophilic granulocytes. Moreover, position -38 is located in the proximity of the binding site for the nuclear factor of activated T-cells (NFAT). Both of these binding sites are important since they affect promoter activity of RNASE3 as the so-called intron-enhancer elements. -38A allele has been associated with interindividual variation in serum ECP levels (31). Also, the carriers of -38A allele more frequently have serum ECP concentration higher than 16 μg/L and develop allergic asthma more frequently than -38C allele carriers.
Population studies of RNASE3 polymorphisms have so far been performed on a small number of subjects, i.e. individual patient groups. For this reason, it is difficult to interpret contradictory results between individual groups observed, and to apply them in routine clinical practice.
As most polymorphisms contribute to phenotype only moderately, multicentric studies of various subject groups, i.e. the genome wide associations (GWA) are necessary in order to reveal the actual causative relationship between SNP and phenotype. A polymorphism may very often contribute to a phenotype only in interaction with other polymorphisms, that is, a certain polymorphism has different effect on the phenotype depending on the degree of exposure of an individual to a particular environmental factor. All this indicates that further studies are yet necessary for better understanding of the role of ECP polymorphism on the development of certain pathological conditions.
ECP and its mechanism of action
ECP exhibits numerous biological activities that may be classified into reactions with cytotoxic effect and those with non-toxic effect.
Cytotoxic activity of ECP is effective against a wide range of microorganisms: parasites, Gram-negative and Gram-positive bacteria, viruses (32-34). Although antimicrobial activity of ECP in in vivo conditions is still intensively investigated, results of in vitro investigations indicate unspecific mechanisms of action (35).
The data gathered are only individual puzzles that are yet to be pieced together into the final mechanism of cytotoxic action.
Antibacterial activity
For antibacterial ECP activity, electrostatic interactions are important between negatively charged cellular membrane or cellular bacterial wall and positively charged ECP, followed by destabilization of bacterial membrane. Other mechanisms involve partial insertion of ECP into lipid bilayers and formation of transmembrane pores that allow the transition of water and osmotic cell lysis. Specific interaction of ECP with lipopolysaccharides and peptidoglycans on the bacterial cell wall may result in bacterial cell aggregation and cell death (35-39).
Antihelminthic activity
The correlation between ECP and antihelminthic activity is based on the information that eosinophilia is present in the peripheral blood of patients with parasitary diseases and granulocyte and eosinophil infiltrates have been detected in tissue biopsy. However, the actual role of ECP and eosinophil granulocytes in parasitary infection has not been clarified. Some studies have suggested that eosinophil granulocytes act on parasites only in their infective larval stages, but not on adult forms (40). As parasitary diseases usually have chronic course, their pathomechanism depicts adaptational processes, that is, tissue damages are not the consequence of action of a pathogenic agent itself but rather occur due to host immune response. One of the forms of such immune response is isolation of pathogenic agent and the entire infected region in the form of granuloma. Blood cells, particularly eosinophil granulocytes, are part of the structure of such granulomas. Over time, some patients develop fibrotic plaques, with ECP as a mediator in their formation. The mechanism by which ECP mediates the remodeling of the extracellular matrix is initiated by the chemotactic action of ECP on fibroblasts. In addition, ECP stimulates the excretion of a pro-fibrotic mediator from fibroblasts, the tumor growth factor-β (TGF-β) (41).
Antiviral activity
Antiviral action of ECP is mediated by ribonuclease activity. Actually, ECP belongs to the family of ribonucleases A (RNase A) which, as exo- and/or endonuclease, cleave the single-strand RNA molecules. Ribonuclease activity of the ECP molecule is considerably lower than the EDN molecule (34,42). Ribonuclease activity is, however, the same in all genetic and posttranslational ECP variants and is entirely independent of cytotoxic activity. The largest number of studies of the mechanism of antiviral ECP action have been performed on respiratory syncytial virus (RSV) because of its size (15.2 kb) and structure (single-stranded RNA genome) (43,44). Results of in vitro investigations showed that ECP lowers viral infectivity, yet ribonuclease activity itself - although important, - is not sufficient for full antiviral effect that is attributed to eosinophil granulocytes (42).
Because of its anti-infective properties, ECP has possible therapeutic potential in the drug development industry. ECP-peptid (1-45) retains the ECP native protein cytotoxicity against bacteria and membrane disrupting activities, too. Furthemore, ECP-peptid (1-45) fulfils the common criteria for antimicrobial peptide as alternative antibiotics (45,46). Understanding specific interactions between pathogenic agents and antimicrobial proteins could be aid in designing new antimicrobial drugs, particularly at in these times of increasing resistance against antibiotics.
Antihost activity
In addition to its important role in host immune defense, ECP may via its cytotoxic activity also cause undesired side-effects on the host’s own tissues. Neuronal damage (Gordon’s phenomenon) has been described, as well as the damage of muscular cells and respiratory tract epithelial cells. If the effect of ECP could be manifested on eukaryotic cells, specific binding of the ECP molecule on the cell surface of heparan sulfate proteoglycans would be of crucial importance. One of the possible mechanisms of eosinophil-induced tissue destruction is based on the apoptotic action of ECP via activation of the caspase cycle (35,47-49). In contrast, intracellular accumulation of ECP molecules is critical for dermal cell damage in numerous dermatoses, while the pathomechanism of skin lesions is based on RNase activity and cation-dependent cytotoxicity of the ECP molecule (50).
Non-toxic activity
Non-toxic action of ECP involves numerous immunomodulatory activities and some authors advocate the hypothesis that the primary in vivorole of the ECP molecule is exactly the general host defense. Some immunomodulatory activities are inhibition of T-cell proliferation, upregulation of receptors and adhesion molecules on epithelial cells, or basophil histamine release (2,35,41). Specific region of ECP action is tissue remodeling which has been particularly studied on the model of asthma. ECP action in tissue repair processes begins with chemotactic action on fibroblasts. Moreover, ECP stimulates the excretion of TGF-β, a factor with pro-fibrotic action whose activity is manifested in altering the intracellular metabolism of fibroblasts. In addition to enhanced proteoglycan synthesis, ECP also mediates the inhibition of proteoglycan degradation, with resulting intracellular proteoglycan accumulation (41,51). Cardiology is a new branch of medicine that has, since not long ago, been dealing with the consequences of ECP activity on atherogenesis. Thus, ECP enables adhesion of monocytes on endothelial cells, is involved in coagulation cascade, and has a stabilizing impact on the plaque (2, 41, 51).
Clinical significance of ECP
Only activated eosinophil granulocytes release the granule content and therefore the determination of ECP concentration is a considerably more specific indicator of eosinophil inflammation that eosinophil granulocyte count in peripheral blood. ECP has been associated with several pathologic conditions. The largest number of clinical studies deal with atopic diseases: asthma, allergic rhinitis, atopic eczema/dermatitis syndrome (AEDS) (53-56). Elevated serum values are proportional to the intensity of allergic inflammation and indicate acute allergen exposure. In addition, diagnostic and prognostic role of ECP is investigated also in other eosinophil-related diseases like infective diseases, respiratory disorders, inflammatory intestinal diseases, malignancies with eosinophilia (27,57-61).
ECP and asthma
ECP has been intensively studied as a potential marker of asthma since the secretory activity of eosinophil granulocytes, particularly the release of degranulation products, is an important component of asthma pathophysiology. Basic proteins from eosinophil granules induce tissue damage, airway inflammation and airway remodeling. However, ECP has no impact on the release of bronchoactive mediators and airway hyper-responsiveness (62,63).
Serum ECP level in asthmatic patients correlates with the biopsy finding of eosinophil granulocytes in the airway mucosa and with the clinical data. Although a large number of studies have confirmed the association of ECP with asthma, ECP determination is not a suitable diagnostic marker of asthma because changes in serum ECP level have been registered in other atopic diseases like allergic rhinitis, but also in conditions not related to the eosinophil inflammation like bacterial sinusitis (54,56). Moreover, eosinophil granulocytes and ECP are parameters whose application is limited only to the allergic type of asthma.
However, as ECP concentration in tissue and body fluids correlates with disease activity, ECP may aid in assessing the severity of asthma and in reaching a decision on therapy type and intensity (54,55), particularly in children because of frequent discrepancy between physician’s assessment and the child’s subjective experience of disease. By following-up changes in the ECP level after introduction of therapy, it is possible to monitor efficiency of a specific drug dose and to correct dosage to avoid overdosing or asthma attack because of a too low dose, all with a view to maintain the well controlled asthma status.
Increasing serum ECP concentration during acute exacerbations is also of clinical significance (55). Thus, the elevation of ECP levels in asthmatic patients with inadequate therapy points to relapse and acute exacerbation, and ECP determination may be used as an indicator of whether patients take their therapy regularly. However, exacerbation may occur also without an increase in serum ECP concentration in some asthmatic patients at an advanced age or in patients with prolonged disease duration. In contrast to this, elevated ECP level have been registered in virus-induced exacerbation, with normal eosinophil granulocyte count in peripheral blood (64).
Measurement of eosinophil activity during the youngest years may aid in predicting the development of asthma at school age. Thus, the children who had been hospitalized at their earliest age due to wheezing in respiratory infection and whose serum ECP level was higher than 20 μg/L had considerably increased risk for the incidence of chronic asthma at school age (specificity 93%, sensitivity 30%) (65).
ECP and hypereosinophilia
Hypereosinophilia is a common laboratory finding that may occur as a consequence of reactive eosinophilia (tissue parasitosis, inflammatory or allergic conditions, malignancy), clonal eosinophilia (hematologic malignancy) or because of unknown cause (idiopathic hypereosinophilic syndrome, IHES). The number of eosinophilic granulocytes in peripheral blood reflects current balance status among cells released from the bone marrow and tissue cells. However, the number of eosinophil granulocytes does not reveal anything regarding tissue infiltration and eosinophil activity. Determination of plasma ECP concentration and the ECP/eosinophil count ratio may promote differential diagnostics of hypereosinophilia. It has been observed that the values of ECP and of the above ratio were the highest in patients with idiopathic hypereosinophilic syndrome (IHES), and that these valus were higher in patients with reactive eosinophilia with malignancy than in patients with reactive eosinophilia with inflammation. However, it is not possible to distinguish clonal from reactive eosinophilia on the basis of ECP concentration and ECP/eosinophil count ratio (59).
ECP and biological specimens
ECP is present in numerous body fluids like plasma, serum, sputum, bronchoalveolar lavage (BAL), saliva, nasal lavage, tears, jejunal fluid, faeces, synovial fluid (2). However, the availability of biological material, invasive sampling methods and complex sample management prior to ECP level determination are the reasons why serum is most commonly used in routine practice.
ECP in serum and plasma
ECP level in serum is higher than its level in plasma because anticoagulants inactivate eosinophil granulocytes and block degranulation ex vivo (66,67). The level of ECP in plasma corresponds to the ECP level in circulation during blood sampling time, i.e. it represents the difference between the ECP released in circulation and the ECP eliminated from circulation.
In circulation, ECP occurs as a free protein and in complex. ECP is bound to α2-macroglobulin (α2-M), and the half-life of ECP in circulation is approximately 45 minutes (68). In addition to being the circulating level, the serum level of ECP represents the secretory activity of eosinophil granulocytes during blood coagulation, and therefore serum ECP concentration (2-20 μg/L) is up to tenfold higher than its concentration in plasma. Serum ECP level allows easier differentiation between healthy and sick subjects in comparison to plasma ECP concentration because the effect of the turnover of ECP in vivo is eliminated in serum. However, in order to ensure correct interpretation of serum ECP concentration, it is necessary to collect the sample for serum ECP determination under standardized conditions (67,69,70). Actually, the type of collection test-tube, temperature and duration of blood clotting, centrifugation and hemolysis are the factors that may cause false positive serum ECP results (Table 1). It is therefore necessary to read properly the instructions of the reagent manufacturer prior to sampling, and to apply the recommended protocol.
Table 1. Effect of sample handling on the eosinophil cationic protein result in serum
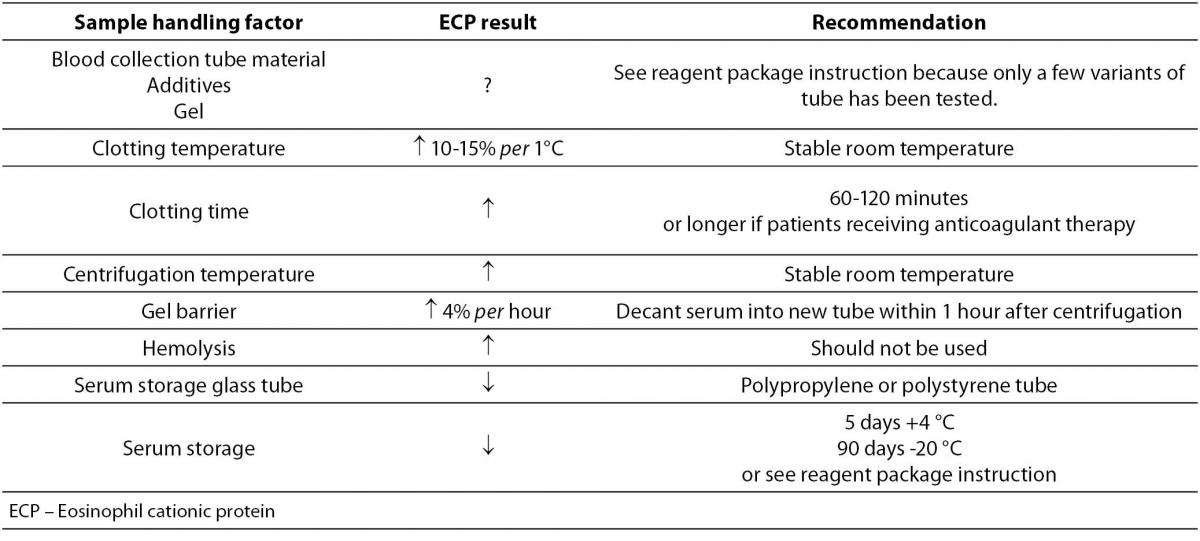
Serum ECP concentration in healthy individuals is characterized by considerable intraindividual variation, 27% (71). During 24 hours, its values are higher at night while the peak serum ECP level is in the morning at 6 o’clock (72). To decrease the impact of circadian variations on serum ECP concentration, it is, for longitudinal comparison, recommended that blood sampling is performed at approximately the same time of the day. In addition to daily changes, seasonal differences in the ECP concentration have also been described in non-atopic children (73). Moreover, gene polymorphisms that change the level of RNASE3 expression are also responsible for interindividual differences in serum ECP concentration (31).
Sputum ECP
Determination of ECP concentration in sputum provides exact information on the local inflammatory process in patients with asthma, chronic obstructive pulmonary disease (COPD) or cystic fibrosis (60,74-77). Moreover, ECP concentraton in sputum (20-1280 μg/L) is considerably higher than that in serum. However, the use of sputum as a routinely applied biological material is associated with some difficulties: the sampling of spontaneously produced or induced sputum, preanalytical sputum management to separate cells from other contents, the effect of saliva contamination, overlapping of normal and pathological ranges of ECP levels (54,78). Therefore, sputum sampling should be standardized before it is used in routine practice (79).
ECP in saliva
ECP concentration in saliva is 250-450 μg/L (54). Saliva sampling is a simple and noninvasive procedure, readily acceptable to children. Correlation between serum ECP and saliva ECP concentration is not confirmed (80). For the routine ECP determination in saliva it is necessary to standardize the sampling method and investigate the clinical utility of such laboratory result (54).
ECP in lavage fluids
ECP concentration in nasal lavage fluid and bronchoalveolar lavage fluid is very low and the collection of the biological material involves invasive sampling methods (54). Moreover, nasal lavage ECP levels are characterized by high intraindividual variation, 123% (81). A novel method to collect nasal secretion by nasal secretion collector shows numerous advantages over nasal lavage method (82). This method could be useful tool for the analysis of nasal inflammation and for following diagnostic and therapeutic intervention (83,84).
ECP in human stool samples
Fecal ECP is intensively investigated as a novel potential marker of inflammatory bowel disease (IBD) and eosinophil gastroenteritis (57,61). Past IBD diagnostics was based on conventional histology which demonstrated the presence of eosinophilic granulocytes in intestinal mucosa; this, however, involved an invasive method of the biological material sampling, whereas fecal ECP determination is a noninvasive and cheaper diagnostic method. Moreover, fecal ECP levels are not dependent on the number of eosinophil granulocytes or serum ECP values, which makes this type of measurement a potential intestinal marker. Also, fecal ECP levels do not differ in dependence to age or gender (85).
Conclusion
The ECP molecule participates in a large number of biological reactions, which makes it an unacceptable diagnostic marker due to low diagnostic specificity. Despite the provision of a large amount of useful data (search term ECP, a total of 2196 references were identified from Pub Med on April 14th 2011), all to date clinical or molecular studies have been associated with certain limitations: small number of subjects (studies ranged from fewer than 10 participants to more than 700 participants), insufficient integration of all factors that may affect interpretation of results, various protocols for preparation of samples for analysis, different analytical methods for ECP level determination, and many others. In addition, many conclusions have been reached on the basis of a very simplified approach where different pathologies are simplified to the level of diseases caused by only one type of cells, eosinophil granulocytes, and a single molecule, ECP. It is presently known that the frequency of eosinophil granulocytes and ECP concentration vary in the same pathological condition in some patients, and that ECP expression is dependent on genetic and posttranslational factors. Therefore, new studies should be designed so as to provide answers to all open questions.